


Introduction
Chronic kidney disease (CKD) refers to a progressive impairment of renal structures and functions. Some studies have reported on the crosstalk between the kidneys and other organs [1,2]. CKD adversely affects brain and nervous system functions [3,4]. However, there is still no consensus on the exact mechanism by which this crosstalk takes place between the kidneys and the nervous system [3,4].
Indoxyl sulfate (IS) is a protein-bound uremic toxin that hemodialysis cannot eliminate [5]. It causes nephrotoxicity, especially in tubular cells, and inhibits endothelial cell proliferation. IS reduction could delay CKD progression. Previous clinical studies have produced no conclusive evidence showing AST-120 (an oral spherical activated carbon) has benefits in delaying CKD progression [6–8]. A recent meta-analysis has shown that tailored AST-120 dosing represents an optimal treatment strategy because it lowers the rates of composite renal outcomes and end-stage kidney disease [9]. In most studies, the primary outcomes are CKD progression and death. The effects of IS reduction on other organs, e.g., the brain, are lacking. Recent studies suggest that IS affects glial function and neuroinflammation in CKD [10,11]. Further studies of chronic IS exposure on glial cells may reveal potential therapeutic targets for cognitive and emotional changes associated with CKD.
Astrocytes provide trophic support for neurons and promote formation and function of synapses [12]. Astrocytes are required for neuronal survival, and the loss of normal astrocyte function can be a primary contributor to neurodegeneration [13]. Several studies report that IS induces apoptosis in human astrocytes through oxidative stress. Especially, a recent study using next-generation sequencing in human astrocytes has revealed that IS can induce pathways associated with oxidative stress, nuclear factor erythroid 2-related factor 2, and mitogen-activated protein kinase signaling pathways [14]. This suggests that IS could be related to CKD-induced neurodegenerative disease [10,14–16]. Furthermore, the mitochondrial central carbon metabolism in astrocytes regulates overall brain bioenergetics, neurotransmitter homeostasis, and redox balances. We recently showed that impaired carbon metabolism, particularly aerobic glycolysis, may be a novel mechanism of Alzheimer’s disease by damaging astrocytes [17,18]. However, the effect of IS on astrocyte glycolysis have not been studied. Consequently, we investigated whether IS inhibits glycolysis and induces apoptosis in astrocytes by reducing glycolysis.
Methods
Cell cultures
The astrocytes used in our study are primary cells isolated from the human brain cerebral cortex (#1800; ScienCell Research Laboratories). The culture method followed the manufacturer’s protocol. GlutaMAX media (10569010; Gibco) with 1% N-2 supplement (17502048; Gibco), 10% fetal bovine serum (26140079; Gibco), and 1% antibiotic (30-002-CI; Corning) was added and 80% confluence cells were cultured in a 100-mm cell culture dish (20100; SPL Life Sciences). Cells were incubated in 5% CO2 at 37 ℃ conditions.
Cell viability
To evaluate the cytotoxicity of IS, we performed a thiazolyl blue tetrazolium bromide (MTT) assay. We seeded 5 × 103 cells per well in a 96-well plate and incubated them for 24 hours. The IS was diluted and processed by concentrating in in a complete media (N-2 supplement with fetal bovine serum). After 7 days, the culture medium was removed, and 100 μL of MTT (M2128; Sigma-Aldrich) at a concentration of 0.5 mg/mL was added per well and incubated for 4 hours. Then, we removed the supernatant and add 100 μL of dimethyl sulfoxide to completely dissolve the generated formazan. Absorbance was measured at 570 nm using a plate reader (Multiskan Go; Thermo Fisher Scientific).
Annexin V and propidium iodide staining
We used annexin V propidium iodide kit (556547; BD Pharmingen) to evaluate apoptosis/necrosis. For confocal imaging, cells were seeded on a sterilized cover glass coated with 2 μg/cm2 poly-L-lysine solution (P4707; Sigma-Aldrich). After 24 hours of incubation, the IS was treated by diluting each concentration. Seven days later, annexin V (Alexa 488-conjugated) and propidium iodide (red-fluorescent) were diluted in the binding buffer provided at 1:100, treated and incubated at 37 ℃ in the dark for 15 minutes. After that, the staining solution was removed, and it was fixed with 4% paraformaldehyde for 5 minutes. Then, it was washed with phosphate buffered saline (PBS) and mounted with FluoroShield with DAPI (F6057; Sigma-Aldrich). The stained apoptosis/necrosis cells were also imaged with a confocal microscope (LSM710; Carl Zeiss).
Fluorescence-activated cell sorting
Before fluorescence-activated cell sorting (FACS) analysis, cells were trypsinized and immuno-labeled with annexin V and propidium iodide. The cells were incubated in a buffer with fluorescent dyes under the manufacturer’s instructions. Then, they were analyzed using a FACSCalibur flow cytometer (BD FACSCanto II; BD Biosciences). The cells with fluorescein isothiocyanate (FITC) annexin V positive and propidium iodide negative were considered to undergo apoptosis.
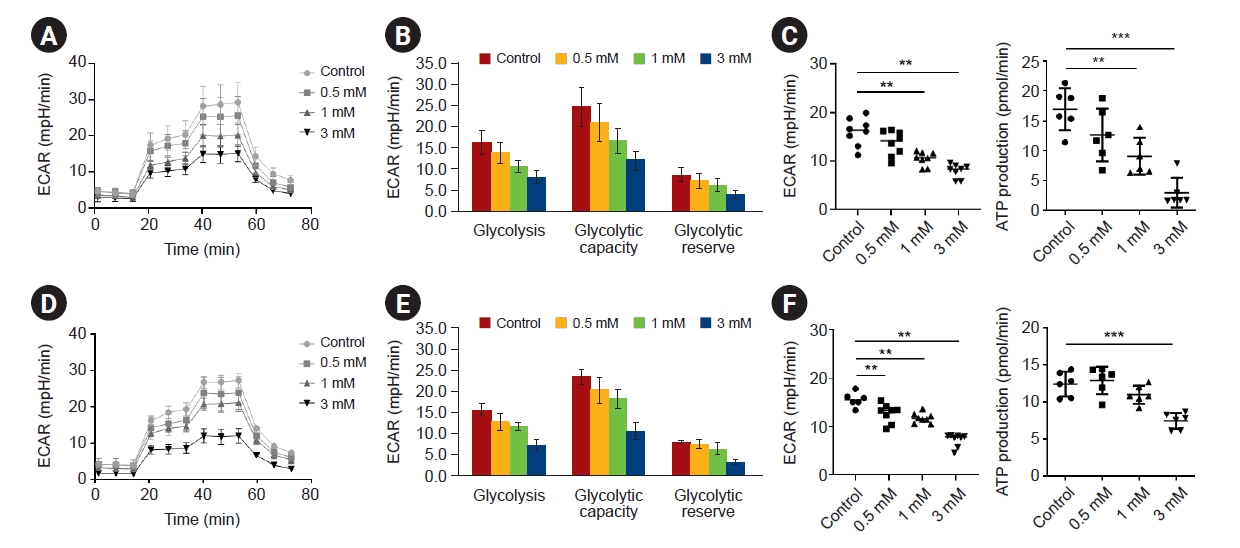
Glycolysis rate assay
To evaluate the effect of IS on the glycolytic function of astrocytes, we used a direct analysis of the concentration of dissociated hydrogen ions by measuring the extracellular acidification rate (ECAR) with an Agilent Seahorse XFe/XF analyzer. The XF Glycolysis Stress Test kit (103020-100; Agilent Technologies, Inc.) was used, and all experimental procedures followed the manufacturer’s user guide. The detailed methods are those referred to in our previous study [17,18]. Shortly, cells were seeded at 5 × 103 cells per well in the plate included with the kit (102601-100, FluxPak mini for XF96 & Xfe96 Analyzers; Agilent Technologies, Inc.), and treated with IS by concentration. After 4 or 7 days, cells were washed twice with PBS, and the culture medium was replaced with XF DMEM Base Medium (103575-100; Agilent Technologies, Inc.), and incubated for 1 hour in a non-CO2 incubator. After that, glucose (final concentration, 10 mM), oligomycin (final concentration, 1 μM), and 2-deoxyglucose (2-DG; final concentration, 50 mM) were added in cartridge ports A, B, and C, respectively. Then, the hydrated cartridge sensor was installed to the cell plate and the assay was performed.
Western blotting
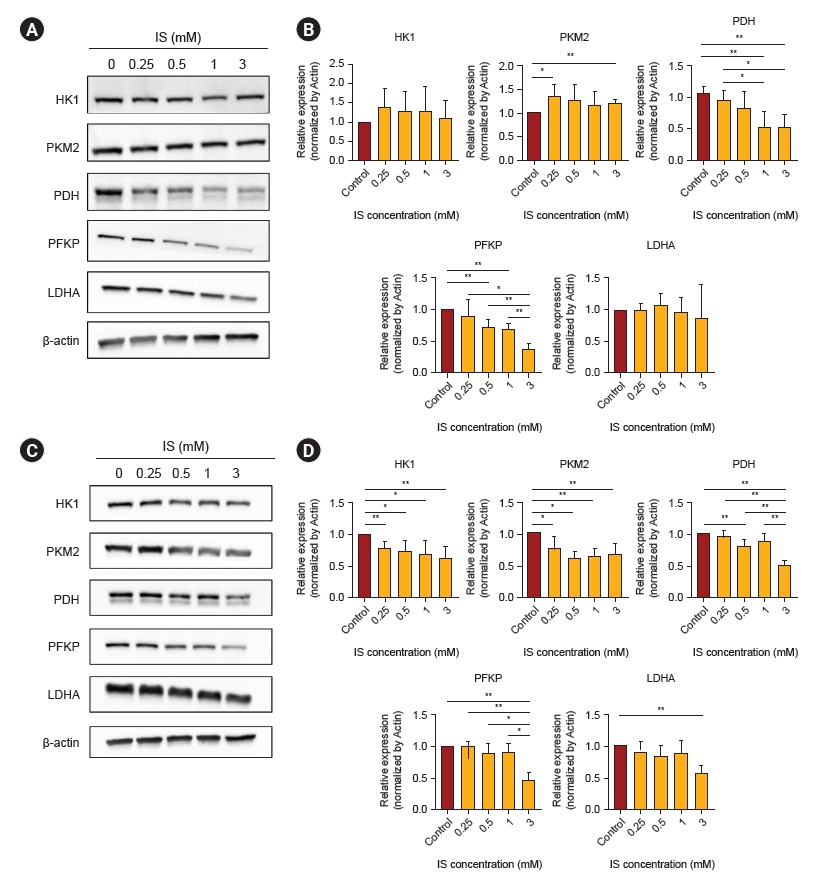
To quantify intracellular proteins, we used western blotting. Astrocytes were seeded 5 × 104 per well into a 12-well plate and incubated at 37 °C and for 24 hours. After that, IS was treated for each specified concentration and time. Then, the trypsinized cell pellet was lysed with radioimmunoprecipitation assay (RIPA) lysis and extraction buffer (89900; Thermo Fisher Scientific) on ice for 30 minutes. After centrifuging at 11,000 rpm for 5 minutes, the supernatant was used for immunoblotting. For protein quantification, Bradford protein assay (5000006; Bio-Rad Laboratories) was used according to the manufacturer’s instructions. Then, 30 μg of quantified protein was separated by size with an electrophoresis system (1658004; Bio-Rad Laboratories) using 10% gel (4561035; Bio-Rad Laboratories). This was then transferred to a polyvinylidene fluoride membrane (1620174; Bio-Rad Laboratories) and allowed to react in blocking buffer (5% skim milk in TBST) for 1 hour. Primary antibodies were added and incubated overnight in a cold room. We purchased antibodies for hexokinase 1 (HK1), pyruvate kinase isozyme M2 (PKM2), pyruvate dehydrogenase (PDH), phosphofructokinase (PFKP), and lactate dehydrogenase (LDH), which were included in the cell signaling glycolysis sampler kit (8337T; Cell Signaling Technology). Antibodies for Bcl2 (4223T; Cell Signaling Technology), Bax (89477S; Cell Signaling Technology) and β-actin (3700S; Cell Signaling Technology) were used. All processes were followed manufacturer’s instructions. After washing three times with TBST for 5 minutes each, the HRP-linked secondary antibodies were diluted 1:2,000 and allowed to react for 30 minutes. They then reacted with the enhanced chemiluminescence solution (RPN2106; GE healthcare) and were detected with an imaging system (FUSION Solo S; Vilber). Each membrane was compared with β-actin as a control, and ImageJ software was used for quantitative analysis.
Cell three-dimensional imaging
We imaged morphological changes of astrocytes with a three-dimensional (3D) cell imaging machine (3D Explore; Nanolive) as in our previous studies [17,18]. Briefly, astrocytes were seeded into a FluoroDish (FD35-100; World Precision Instruments) and incubated. Except for the IS treatment, after 24 hours, the astrocytes were treated with IS for 7 days, and those in a control group were incubated under the same conditions. Then, 3D images of astrocytes from both groups were acquired.
Results
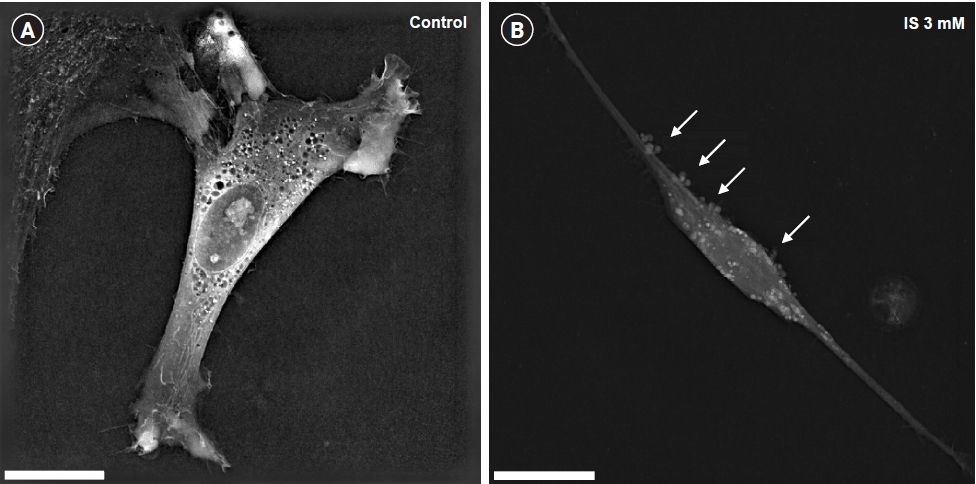
Indoxyl sulfate induced cytotoxicity in human astrocytes
We analyzed the morphological changes of cytotoxicity following IS treatment (at 7 days) in human astrocytes using a 3D cell analyzer. In the IS-treated astrocytes, we observed such apoptotic cell morphological changes as blebbing and shrinkage of the cytosolic area as compared to the control group (Fig. 1).
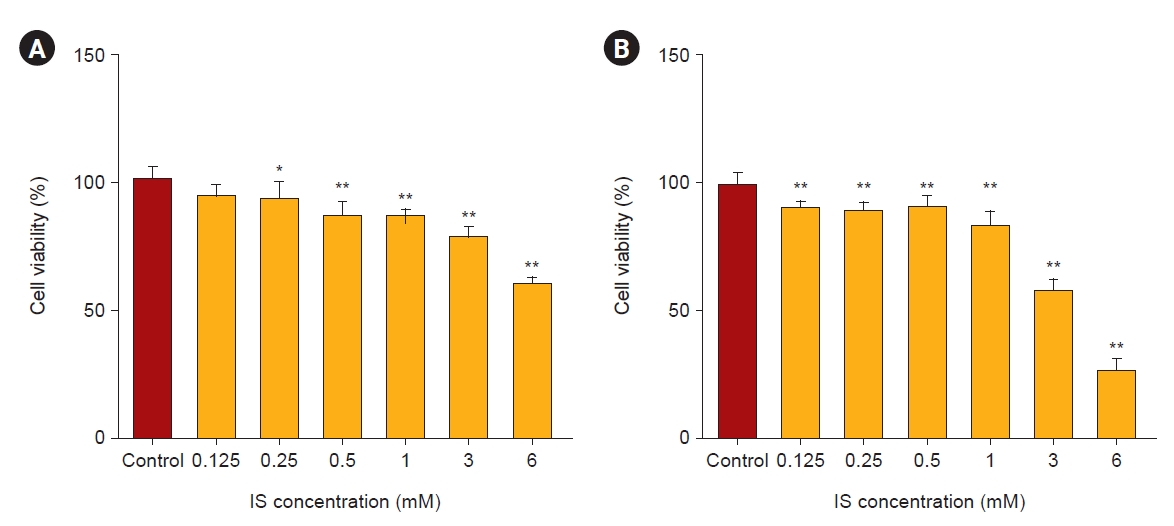
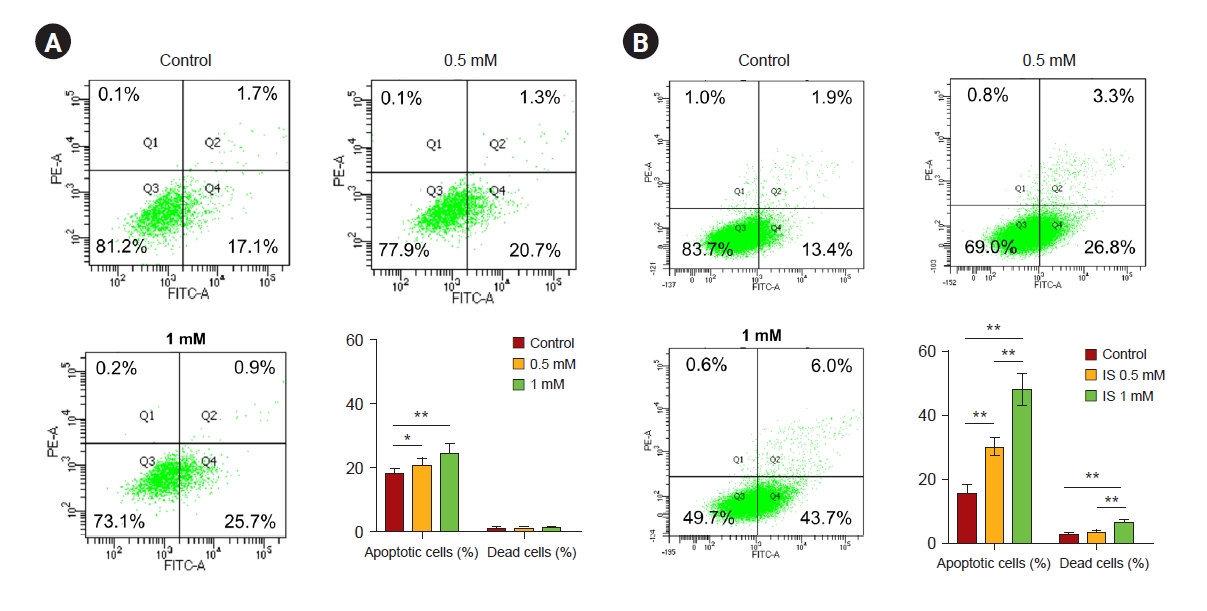
Indoxyl sulfate induced apoptotic cell death in human astrocytes
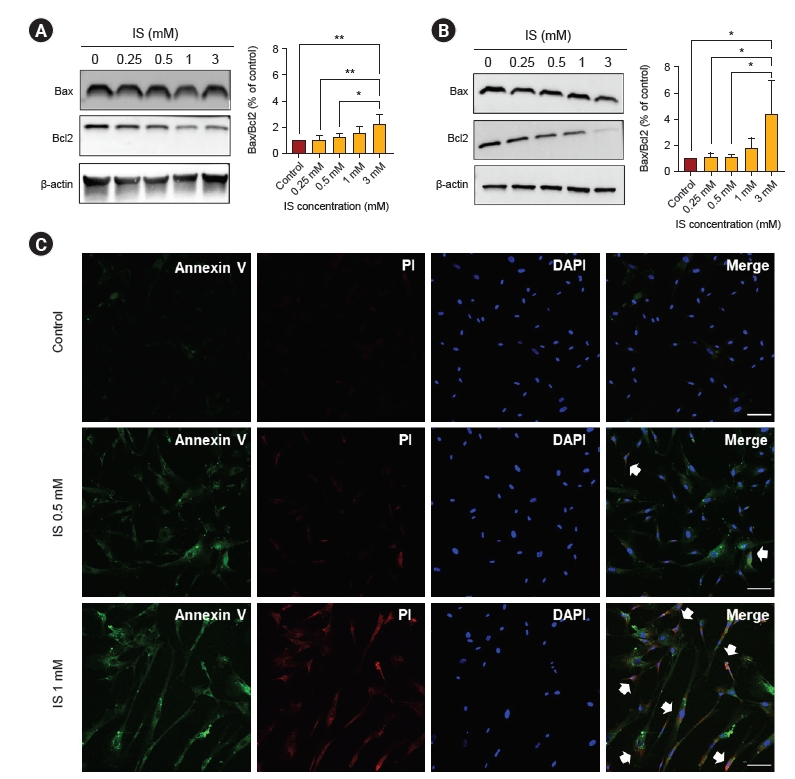
We performed an MTT assay to evaluate the viability of astrocytes as a marker of cytotoxicity by IS. As shown in Fig. 2, after 4 and 7 days, IS reduced astrocyte cell viability in a dose-dependent manner. In order to investigate whether this decreased cell viability was induced by apoptosis as shown in Fig. 1, we did a FACS study. After incubation with IS for 4 and 7 days, it was noted that the proportion of apoptotic cells had significantly increased (Fig. 3). Apoptotic cells were more prominent after 7 days of IS incubation as compared to those after 4 days. This effect was also more noticeable at higher IS dose. An immunoblot study confirmed that the Bax/Bcl-2 ratio had significantly increased by days 4 and 7 in a dose-dependent manner (Fig. 4A, B). Astrocytes incubated both with or without IS (0.5 and 1.0 mM) for 7 days were stained with annexin V, FITC (green), and propidium iodide (red). As shown in Fig. 4C, astrocytes without IS were not stained by either annexin V or propidium iodide. However, cells treated with 0.5 mM of IS were partly stained by either of these and those treated with 1.0 mM were significantly stained. Thus, we were able to readily observe co-staining in these spots.
Indoxyl sulfate suppresses the activation of glycolysis in human astrocytes
We investigated whether IS could affect glycolytic activity in human astrocytes. ECARs were investigated as surrogates of glycolysis by quantifying lactate production. The ECAR was measured during the sequential addition of glucose (the substrate of glycolysis), oligomycin (a selective inhibitor of mitochondrial respiration), and 2-DG (a specific inhibitor of glycolysis). IS significantly suppressed ECAR levels in response to glucose as compared to controls on days 4 and 7, and the degree of suppression was dose-dependent (Fig. 5). Decreased adenosine triphosphate (ATP) levels were also observed.
Indoxyl sulfate reduces the expression of glycolytic enzymes in human astrocytes
Next, we investigated whether IS could regulate enzyme expression in the glycolysis pathway. On day 4, IS decreased the PDH and PFKP protein levels as compared to the control, but the PKM2 seemed to increase slightly (Fig. 6A, B). After 7 days of IS treatment, HK1, PKM2, PDH, and PFKP protein levels were decreased as compared to the control (Fig. 6C, D). These results suggest that glycolysis inhibition in human astrocytes is associated with the exposure time and dose of IS.
Discussion
Our study showed that long-term exposure to uremic toxins suppresses glycolysis in astrocytes. Our results also suggest that the inhibition of glycolysis by IS in astrocytes leads to cell death via apoptosis. IS affects glycolysis in astrocytes not only dose-dependently, but also time-dependently. Specifically, long-term and higher-dose exposures had more serious effects on astrocytes. Because CKD patients are inevitably subjected to long-term exposure to uremic toxins during their disease progression, even slight increases in concentration of uremic toxins can inhibit glycolysis and reduce the activity of astrocytes.
Neurologic disorders, such as Alzheimer’s and Parkinson’s disease, are associated with impaired brain energy metabolism [19,20]. The significance of aerobic glycolysis in astrocytes has been extensively documented in metabolic brain diseases, particularly in Alzheimer disease, where it is known to play a crucial role in its pathogenesis [21,22]. Because of role of glucose as the main energy source in astrocytes, it plays a crucial role in the central nerve system by regulating synaptic activity and intellectual disability [7], with glycolysis and lactate production being metabolic features of astrocytes [23,24]. Indeed, we observed an association between memory impairment and suppressed glucose uptake in the human brain, which we demonstrated with an F-18 fluorodeoxyglucose positron emission tomography scan [25]. In contrast, neurons meet their energy demands predominantly by oxidative phosphorylation [26,27]. Thus, aerobic glycolysis, an energy-producing pathway that uses glucose as metabolic substrates under conditions of sufficient oxygen supply, is essential for memory formation and consolidation [26]. However, whether impaired energy production in the brain contributes to cognitive dysfunction in patients with CKD remains unclear.
Current clinical and animal studies suggest that CKD leads to cognitive impairment and emotional changes [3,11,28]. Previous studies have shown that IS may play a key role in the neurologic consequence of kidney failure [11,29]. The mechanism of how IS affects astrocytes has not been fully demonstrated. A previous study has shown that IS induces human astrocytic apoptosis under oxidative stress by inhibition of the mitogen-activated protein kinase pathway [14]. A damaged blood-brain barrier is associated with cognitive dysfunction [30]. In our recent work, we observed blood-brain barrier disruption in a rat CKD model [31]. Aryl hydrocarbon receptor activation by IS is associated with blood-brain barrier disruption, leading to cognitive impairment in animal CKD models [15].
Sato et al. [32] have demonstrated that exposure to IS can inhibit glycolytic activity in C2C12 myotube cells, which can be partially restored after 24 hours. They suggested that this compensatory regulation of glycolysis resulted in an excess antioxidative response, such as the pentose phosphate pathway [32]. In contrast, our study found that a 4-day exposure to IS suppressed PDH, while a 7-day exposure suppressed further steps in the glycolysis pathway, suggesting a more severe inhibition of glycolysis with prolonged exposure duration. The PDH complex irreversibly converts pyruvate and oxidized nicotinamide adenine dinucleotide (NAD+) into acetyl-CoA, nicotinamide adenine dinucleotide-reduced (NADH), and carbon dioxide [33]. PDH activity must be finely regulated to maintain cellular energy homeostasis and to supply the necessary carbon for biosynthetic pathways that cross the citric acid cycle [33]. Symptoms of PDH deficiency include lactic acidosis, elevated pyruvate levels, and ataxia, along with developmental delay, psychomotor retardation, and decreased cognitive capacity in long-lived patients [34]. Prolonged exposure to IS (7 days vs. 4 days) also suppressed glycolysis more acutely, suggesting that prolonged IS exposure inhibits astrocyte function more strongly. This may be a novel mechanism for cognitive impairment of CKD patients, in which the cognitive function slowly and chronically declines.
Our study has two other clinical implications. First, it can be presumed that long-term exposure to uremic toxins induces brain energy deficiency, leading to cognitive impairment. Thus, improving brain metabolism could be a novel therapeutic option. Glucagon-like peptide-1 (GLP-1) is an intestinal L-cell-derived incretin hormone that stimulates insulin secretion in the beta cells and inhibits alpha cell glucagon secretion of the pancreatic islets [35]. GLP-1 receptor agonist is associated with a lower risk of kidney and cardiovascular outcomes in type-2 diabetes [36,37]. Moreover, GLP-1 agonist improves cognition in 4-month-old five mice with familial Alzheimer disease mutation by enhancing aerobic glycolysis and reducing oxidative phosphorylation levels and oxidative stress in the brain [38]. We hope that further study could reveal whether enhancing glycolysis may improve cognition in CKD. Second, astrocytes contribute to the protection and survival of neurons. The study of astrocytes is particularly important, given the coexistence of neuronal and astrocytic apoptosis in damaged brains suffering from ischemia and neurodegenerative diseases [39]. IS is associated with glial cell neuroinflammation [10,40], and our study demonstrates that IS induces apoptosis in astrocytes. Further studies are needed to reveal the functions and interactions between glial cells and astrocytes in uremic conditions and their role in neurologic complications in CKD patients.
In conclusion, our study shows that prolonged exposure to IS significantly suppresses glycolysis in astrocytes, which culminates in apoptosis. This could be a novel therapeutic target and further studies are needed to estimate whether improvement in brain metabolism could prevent neurologic complications in CKD patients.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print




")









